抗菌肽
抗微生物肽(Antimicrobial Peptides, AMPs)是生物体免疫系统在受到异源刺激时,诱导产生的一类防御性生物活性肽,是机体非特异性天然防御系统的重要组成部分,广泛存在于自然界各种生物以及人体内。
抗菌肽概念的发展
人们对抗菌肽的关注始于20世纪70年代。1975年,瑞典化学家Boman从惜古比天蚕(Hyalophora cecropia)蚕蛹中提取到一种具有抗菌活性的肽类物质,并为其命名为 “天蚕素(cecropins)”。随后人们在多种生物体内陆续发现了多种类似的生物活性肽,如蛙皮素(magainins)、蜂毒素(melittins)和防御素(defensins)等。最初,人们发现这类生物活性肽具有广谱高效的杀菌活性,因而又被称作“抗菌肽(Antibacterial peptides,ABP)”。随着对抗菌肽研究的深入,人们发现某些抗菌肽对真菌、原虫、病毒及癌细胞等均具有杀伤作用,并且还可以促进伤口愈合。因此,许多学者又称其为多肽抗生素(Peptides Antibiotics)。在国内,人们还是比较习惯于“抗菌肽”这一名称。抗菌肽的合成与扩散均非常迅速,作用灵活。所以Boman曾指出,抗菌肽是机体的一种理想的一线防御。其实,能杀灭或抑制微生物生长的肽都可以被称作抗菌肽。因此,广义上的抗菌肽还应包括由细菌产生的细菌素 (bacteriocins),如由乳酸菌产生的乳链菌素(nisin)以及由大肠杆菌产生的大肠杆菌素(colicins)等。迄今为止,从不同生物体内诱导的抗菌肽已不下千种。
抗菌肽的分类:
按照抗菌肽的氨基酸组成和结构的不同可分为以下几类:
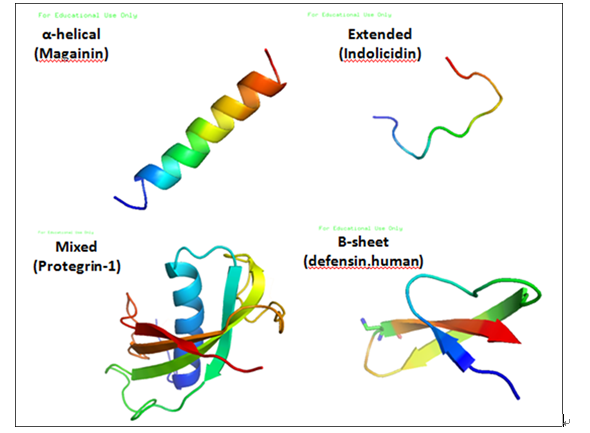
(1)单链无半胱氨酸(Cys),N-端和C-端各有一段α-螺旋,的阳离子多肽。这类抗菌肽在水溶液中常处于无秩序状态,但在脂溶性介质中会形成α-螺旋结构。例如,天蚕素Cecropins和蛙皮素Magainins。Cecropin是分子量大约4 kDa的阳离子肽,由29~42个氨基酸残基组成,是第一个被发现的昆虫抗菌肽。1981年由Boman等人从惜古比天蚕(Hyalophora cecropia)蛹中分离得到cecropin A和B。随后,人们从果蝇、家蚕、柞蚕等昆虫以及线虫甚至猪小肠中都分离出了cecropin。这说明cecropin在于动物界中分布广泛。Cecropins的N-端具有强碱性,富含赖氨酸和精氨酸等阳离子型氨基酸;C-端为疏水区,富含丙氨酸、缬氨酸和甘氨酸等非极性氨基酸。因此可形成双亲螺旋结构。多数Cecropins的C-端被酰胺化,酰胺化对保持其抗菌活性和稳定性具有重要作用。Cecropins对革兰氏阳性菌以及一些革兰氏阴性菌具有很强的杀伤力。目前发现的天蚕素A-F主要发现于鳞翅目和双翅目。天蚕素类还包括了分离自天蚕的cecropinA、B、cecropin D-G,家蚕的cecropinA、B、D,柞蚕的cecropin B、D,黑腹果蝇的cecropin A1,A2、B,棕尾别麻蝇的sarcotoxins以及hyphancin、enbocin和Spodopsin等几十种抗菌肽。Magainins最初是从非洲爪蟾的皮肤中发现的它是爪蟾的皮肤在一定的环境压力下分泌出的抗感染和促迸伤口愈合的成分,低浓度便可抑制许多细菌和真菌生长。研究表明,α-螺旋的含量与抗菌肽对G-菌和G+菌的活性密切相关。
(2)富含某种氨基酸残基但不含Cys的抗菌肽。这类肽一般为线性结构,有的形成延伸的卷。例如,富含Try的牛中性粒细胞多肽indolicidin,富含Pro和Arg的细菌素和猪肠源抗菌肽PR-39,鞘翅肽Coleoptericin和半翅肽Hemiptericin,蜜蜂肽Apidaecins,源于果蝇的Drosocin,人源抗菌肽Histatins还有富含Pro和Phe的prophenin都属于此类。Indolicidin的13个氨基酸中含有 5个色氨酸而得名,其C端是酰胺化的,对大肠杆菌和金黄色葡萄球菌都具有很强的杀菌活性。PR-39的Pro含量占49%; Coleoptericin和Hemiptericin富含Gly。Apidaecins含有 16~18个氨基酸残基,其中Pro含量高达 33%,Arg含量可达 17%。Drosocin富含Pro,在结构上与 apidaecins 具有一定的相似性,但是在其 11 位的Thr羟基上连接着O-二糖链(-N-乙酰半乳糖胺-半乳糖)。 Histatins是从人的腮腺和下颌腺分泌物中分离得到的一组富含His的抗菌肽,长度为 7~38个氨基酸残基不等,对于引起口腔感染的多种微生物具有活性。
(3)含一个二硫键的抗菌肽,该二硫键的位置通常在肽链C-端。此类抗菌肽数量较少,第一个被发现的是来源于牛中性粒细胞的bactenecin,其 12 个氨基酸中含有 4个Arg,在其第2位和第11位氨基酸残基间形成二硫键。Bactenecin 对大肠杆菌和金黄色葡萄球菌都有活性。该类抗菌肽中还包括来源于蛙类皮肤的Brevinins, C-端有一个由7个氨基酸形成的环和一个长的 N端尾巴。
(4)含两个或两个以上由Cys形成的二硫键,具有稳定β-折叠结构的抗菌肽。此类抗菌肽常为环状结构。防御素即属此类。与天蚕素类抗菌肽相比,昆虫防御素在结构上更为一致,不同的防御素蛋白的一级结构具有高度的同源性。从结构上分析,昆虫防御素分子中富含Cys,分子量大约为4~6 kDa,通常由33~46个氨基酸残基组成。从蜜蜂中得到的royalisin,由51个氨基酸残基组成;从斑腹刺肩蝽分离的thanatin,仅有21个氨基酸残基,含2个半胱氨酸,1个二硫键,是该类抗菌肽中最小的一个。绿蝇防御素Phormicin有6个Cys,形成3个分子内二硫键,肽链C-段是带有拟β-转角的反向平行的β片层。猪白细胞肽protegrin含有16个氨基酸,通过4个Cys形成两个二硫键。研究表明,分子中的二硫键在其抗菌作用中至关重要。
(5)由已知功能蛋白衍生而来的抗菌肽。例如,来自于乳铁蛋白、Casocidin I以及来自于α-乳白蛋白、人血红蛋白、溶菌酶和卵清蛋白的抗菌活性片段。这些片段具有抗菌活性并在组成和结构上与其它抗菌肽类似。另外,细菌素常由其前体经翻译后修饰衍生而来。根据衍生情况,可将革兰氏阳性菌抗菌肽分为2类:一类是翻译后经修饰蛋白分子结构中有羊毛巯氨酸基团的抗菌肽,即羊毛巯类抗菌肽lanbiotics类。此类分子成可塑的长链状,通过在生物膜上形成膜孔杀灭细菌。包括乳酸链球菌肽nisin,枯草菌素subtilin,sublancin,表皮素epidermin,Pep5以及变链素mutacin等。其中研究最广泛的是nisin,它来源于乳酸菌,含有羊毛硫氨酸、甲基羊毛硫氨酸等特殊基团。另一类是未修饰的抗菌肽,即非羊毛巯类抗菌肽类,包括mersacidin, actagardine, 肉桂霉素cinnamycin。通过抑制细菌细胞壁肽聚糖的生物合成来杀灭细菌。
(6)阴离子抗菌肽。例如,Maximin H5、Dermcidin以及富含Glu和Asp的阴离子小肽。它们发现于表面活性提取物、支气管肺泡灌洗液和气管上皮细胞。它们以Zn2+作为辅助因子,对G-茵和G+菌都有活性。
抗菌肽的氨基酸组成和结构对其与细胞质膜相互作用,进入细胞质膜、核膜至细胞内作用靶位起作用有重要意义。
抗菌肽的活性
抗菌肽作为一种非特异性免疫因子,具有高效广谱的杀菌活性。某些抗菌肽不仅对细菌有杀伤力,对真菌、病毒、原虫和癌细胞也有杀伤作用,而且还对癌细胞有选择性杀伤作用。
1. 抗细菌活性
抗菌肽可直接快速地杀灭包括高度耐药菌株在内的许多致病菌。抗菌肽的种类繁多,不同的抗菌肽也表现出各自不同的抗菌活性特点。例如drosocin、attacins、deptericins等对革兰氏阴性菌有较强杀伤作用,defensin则对革兰氏阳性菌较为敏感,而果蝇Cecropin既抗革兰氏阳性菌又抗革兰氏阴性菌,甚至对真菌也有活性。同一抗菌肽对不同的菌株的活性也是不同的。由于抗菌肽作用机制有别于传统的抗生素,能杀死耐药性的病原菌突变种,而病原菌却不产生抗药性。
2. 抗真菌活性
随着对抗菌肽研究的逐渐深入,抗菌肽的抗真菌活性陆续被发现。黑腹果蝇的drosomycin是第一个被发现的抗真菌肽。drosomycin仅对丝状真菌有较强的抗性,而对细菌和酵母不敏感。Termicin是一种白蚁抗真菌肽,对丝状真菌和酵母均有活性,由36个氨基酸残基组成,含6个Cys,形成3个二硫键。同样来源于白蚁的spinigerin由25个氨基酸残基组成,不含Cys,呈α-螺旋结构,对丝状真菌和酵母的抗菌效果较强,是一种的线性多肽。现已发现有几十种昆虫多肽有抗真菌活性。抗真菌肽的发现,为治疗真菌感染性疾病提供了新的有力手段。
3. 抗肿瘤细胞活性
研究发现,一些抗菌肽能特异地抑制某些肿瘤细胞生长,而不影响正常体细胞。抗菌肽能够直接定位并作用于体外培养癌细胞的质膜结构,使膜上形成孔洞,内容物外泄。被作用的细胞还会出现线粒体出现空泡化、嵴脱落、核膜界限模糊不清甚至核膜破损,核染色体DNA断裂并可抑制染色体DNA的合成。例如,Cecropin B可通过细胞间接触作用杀死癌细胞。抗菌肽对肿瘤细胞的选择性灭杀、分子量小、热稳定性好、无免疫原性等特点使其很有潜力成为抗肿瘤新药。
4. 抗原虫
抗菌肽对痢疾、疟疾、Chagas氏病、莱什曼病原虫等原虫也有杀伤作用。研究发现,抗菌肽对疟原虫的卵囊期和子孢子期的疟原虫有明显的杀灭作用。可使疟原虫卵囊密度明显降低,内部形成大的空泡;而子孢子的膜通透性受破坏,形态受损,流动性降低。cecropin B人工改造后也可杀死虐原虫。
5. 抗病毒能力
研究表明,蜂毒素和天蚕素可在亚致死浓度下抑制艾滋病毒HIV-1基因的表达,从而减少其增殖。这表明抗菌肽对病毒也有治疗作用。除此以外,抗菌肽对流感病毒、乙型肝炎病毒等也有抗病毒作用。抗菌肽的抗病毒潜能对于抗病毒药物的开发具有深远意义。
6. 促进伤口愈合
正常机体在受到局部创伤、感染后会在局部区域产生抗菌肽,抗茵肽的最早临床研究也是从局部感染治疗开始。例如,源于蛙皮素的MSI-78对糖尿病患者的足溃疡有显著疗效;源于猪Protegrin的IB-367可用于治口腔溃疡;而细菌素可用于治疗胃溃疡等。
抗菌肽的作用机制
抗菌肽的作用广泛,对细菌、真菌、病毒、癌细胞、寄生虫等都有杀死或者抑制作用。来源和分类不一样的抗菌肽其作用方式也各不相同。但几乎每种抗菌肽杀菌时都会造成由细胞膜的损伤而导致的细胞内物质的泄露。人们对抗菌肽的杀菌机理的解释也主要在于抗菌肽破坏病原微生物的细胞膜。这可能是抗菌肽最主要的抗菌机理。而哺乳动物细胞膜中含有大量的膜蛋白和胆固醇,细胞还拥有高度发达的骨架系统,这使的抗菌肽对真核细胞没有毒性作用,从而实现抗菌肽对病原专一作用的优点。随着研究的深入,人们发现抗菌肽还可以其它的方式产生抗菌效果,但其杀菌效果的发挥是多种因素协同的综合结果。例如,抗菌肽在细胞内的作用需以进入细胞内为前提。目前对抗菌肽的作用机制研究较多的有以下几种:
1. 对质膜的作用
早在1984年就有研究报道抗菌肽起到了离子泵的作用,使细胞内K+离子外泄、ATP含量迅速下降,继而导致细胞死亡。1988年有研究者通过测定抗菌肽处理后的脂质体的膜电势和电流变化,判断出抗菌肽在膜上形成了孔道。1993年又发现孔道的形成、开启和关闭都依赖于膜的电势,又将其称为电势依赖性通道。1996年研究人员观察到了天蚕素(Cecropin A)结合到大肠杆菌膜上,并行成病灶,最终导致细菌细胞内容物外泄死亡,从而直观的证明了抗菌提对质膜的作用。抗菌肽的正电荷可通过静电作用与细胞膜外层结构结合。革兰氏阴性菌的细胞外膜含有带负电的脂多糖(1ipopolysaccharide,LPS),抗菌肽可取代结合在LPS上的Mg2+、Ca2+等二价阳离子,引起外膜破裂或穿孔而透过外膜。通过外膜的抗菌肽和细胞内膜上带负电荷的磷脂通过静电引力结合,在细胞膜形成空洞或临时通道,从而引起崩解或透过细胞膜。对此,国内外学者对此研究较多。其作用方式可分为“栅桶(barre—stave)”模型和“毡毯(carpert)”模型。
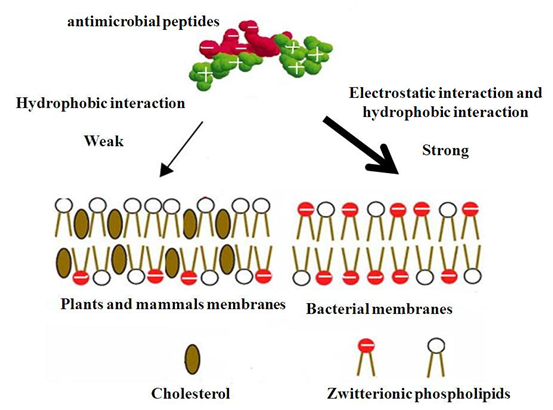
抗菌肽对细菌细胞膜的选择性结合
(1)栅桶(barre-stave)模型:阳离子抗菌肽在电荷的作用下,结合在细胞膜表面,并相互聚合,肽分子中的疏水基团浸入磷脂双分子层内,并与膜表面以垂直方式排列,形成横跨细胞膜的离子通道。离子通道一旦形成,外界的水分即可进入细胞内部,细胞质也可渗透到外部。由于失去能量,严重时细胞膜会崩解而导致细胞死亡。该模型中起主要作用的是阳离子电荷和疏水作用,因此稳定的两亲结构非常重要。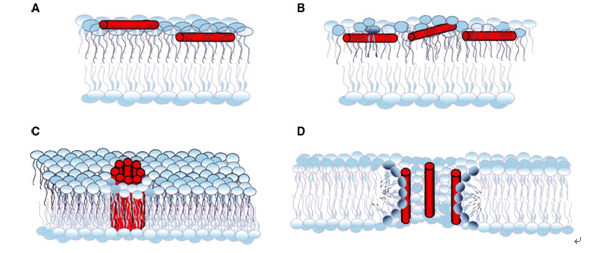
(2) 毡毯(carpert)模型:有些抗菌肽并无稳定的α-螺旋或者β-折叠结构,并且在作用过程中无明显的细胞膜破坏现象,但是仍然可以造成细胞中K+的渗出。此现象可由毡毯模型来解释。该模型认为抗菌肽与细胞膜上的负电荷部结合,平行地排列在细胞壁表面形成类似“毡子”的结构。抗菌肽的疏水部分通过改变细胞膜的流动性和厚度,在细胞膜上出现瞬时空洞,导致细胞质外渗或抗菌肽渗透到细胞质中。这同样会引起细胞膜崩解,导致细胞死亡。该模型其关键作用的是电荷间的作用力,阳离子抗菌肽无需α-螺旋、β-折叠等稳定的二级结构,也不需要在细胞膜表面形成多聚体。抗菌肽引起的细胞膜破坏可以发生在整个细胞膜上,也可仅在其中某些部位。
2 抑制细胞的呼吸作用
有些抗菌肽如死亡素 (Thanatin)对细菌表现出强烈的杀伤作用。但却没有导致细菌细胞内K+离子的外泄。这表明此抗菌肽不是通过改变细胞膜的通透性来杀菌的。研究表明,Thanatin是通过抑制细胞的呼吸作用来杀菌的。
3 抑制细菌蛋白、DNA的合成
有些抗菌肽如Attacins能够干扰细菌细胞外膜蛋白OmpC、OmpF、OmpA以及LamB基因的表达,从而导致细胞膜的通透性增加,细菌的生长受到抑制。Apidaecin抗菌肽与细菌外膜结合,而后进入细胞外周胞质,再通过与细胞内膜的受体分子特异性结合,进入细胞内。最后在胞内抑制蛋白合成,从而起到杀菌效果。Tachyplesin可结合在DNA的小沟,pleurocidin、dermaseptin、PR-39、indolicindin和防御素均可阻止大肠杆菌细胞对胸嘧啶脱氧核苷、尿嘧啶核苷和亮氨酸的摄入从而抑制细菌DNA,RNA和蛋白质的合成。PR-39能够终止敏感细菌DNA复制相关蛋白质的合成。HNP-1和HNP-2能够减少敏感细菌DNA,RNA和蛋白质的合成,其中HNP-1同时还能够抑制细胞周质β-半乳糖苷酶的合成。Indolicidin能够完全抑制E. coli的DNA和RNA的合成,但只有在高浓度下才能够明显的抑制蛋白质的合成。抗菌肽在胞内还可以导致蛋白、DNA的降解。
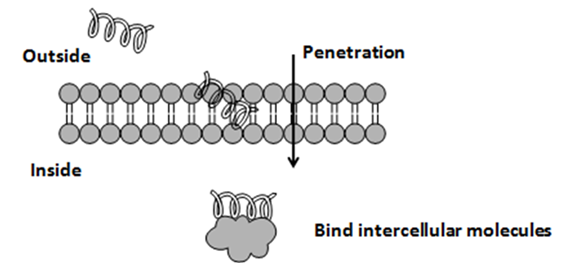
4. 抑制细胞壁的形成
麻蝇毒素II(Sarcotoxins II)的抗菌机理是抑制正在形成的细胞壁,使细菌不能维持正常的细胞形态而生长受阻。mersacidin通过干扰细胞膜相关的转糖基作用而抑制肽聚糖的生物合成;Mersacindin与细胞膜的lipid II结合,而抑制肽聚糖前体转化为聚合的新生肽聚糖。
5. 抗菌肽对真菌以及寄生虫的作用
许多抗菌肽除了具有抗细菌的活性外,还有抗真菌的作用。研究者对由天蚕素A和蜂毒素分子的片段合成的杂合肽分子CA-ME抗真菌的研究发现,在抗菌肽的作用下真菌细胞壁被破坏,细胞也无法保持正常的形态。表明抗菌肽对细胞质膜起作用是由于膜形成孔洞而达到杀菌目的。Histatins可通过结合于真菌细胞膜的特定受体而进入细胞质膜并诱导细胞非裂解模式的ATP损失,与此同时还能够破坏细胞周期并引起细胞活性氧的大量产生。另有研究发现,天蚕素-蜂毒素杂合肽对莱什曼原鞭毛虫的作用靶目标是细胞质膜。作用模式与杀菌模式相似。抗菌肽能快速降低H+/OH-的通透性使细胞的ATP合成下降,呼吸减弱,外膜坍塌,原生质膜形态遭破坏,从而杀死原虫。
6. 抗菌肽对病毒的作用机制
目前研究发现抗菌肽对癌细胞的作用主要是在癌细胞膜上形成孔洞,使其内容物外泄,线粒体出现空泡化,破坏核膜,致使染色体DNA断裂,并抑制染色体DNA的合成,细胞骨架也受到一定程度的损伤。α-defensins、modelin-1等对疱疹病毒的作用以及polyhemusins对HIV病毒的作用是通过抗菌肽直接与病毒离子相结合而实现的。Cecropin A对HIV病毒的作用是抑制病毒的繁殖。Mellitin及其类似物K71的结构与烟草花叶病毒核衣壳mRNA相互作用的区域具有相似性,这样蜂毒蛋白分子就可以伪装成病毒包被蛋白,被病毒RNA结合,导致RNA的构象改变,病毒颗粒无法正常组装,从而通过模仿病毒的侵染过程发挥其抗病毒的作用。此外,抗菌肽还可以调节机体的免疫机能,增强机体的免疫力。
7. 抗菌肽对肿瘤细胞的作用机制
抗菌肽对某些癌细胞有DNA的断裂作用,研究者用单细胞凝胶电泳技术(SCGE)研究了抗菌肽对人髓样白血病细胞和正常人白细胞核染色质的影响,并发型抗菌肽可使癌细胞的细胞核染色质DNA断裂,而正常人的白细胞和未经抗菌肽处理的癌细胞未观察到DNA的断裂。还有研究者发现,抗菌肽首先作用于靶细胞外膜,然后破坏内膜系统即细胞器,最后破坏核膜。由此推测抗菌肽有可能对核内的染色体起作用。
8. 促进免疫作用
抗菌肽能够激活细胞非膜表面的酶,如自溶素和磷脂酶。宿主来源的分泌性磷脂酶A2含有带负电荷的磷脂或卵磷脂,Magainnins、indolicindin、temporins B和temporins L在Ca2+存在的情况下能够显著增强磷脂酶A2对脂质体的水解活性。这种促进的作用很有可能对宿主的天然免疫具有重要意义。
抗菌肽的制备
1. 从天然有机体中提取
抗菌肽的传统分离方法主要是通过人工诱导刺激抗菌肽的产量,再用生物化学方法将抗菌肽从宿主机体中分离出来。例如,昆虫来源的抗菌肽,多是注射抗原后通过收集血淋巴液,再进行分离的。这种传统分离提纯具有抗菌活性的组分是一件非常繁琐的工作。而且不同的试验条件对于组分的效果影响很大。最重要的是受来源的限制。这种以传统方法获得的抗菌肽,不足以满足抗菌肽大规模生产的需要。这也是限制抗菌肽生产成本的主要因素。
2. 化学合成
化学合成法是获得抗菌肽的主要手段之一。通过化学方法可人为设计和控制抗菌肽的序列,是研究抗菌肽结构与功能的有利工具。然而,化学合成法也有其致命的弱点。例如,对于序列较长、结构较为复杂的抗菌肽或者在天然宿主体内合成时有翻译后加工等过程的抗菌肽就不适宜化学合成。成本高也是限制化学合成获得抗菌肽生产的主要原因。
3. 基因工程表达
提高抗菌肽的生产效率、降低成本是抗菌肽应用领域研究中的重要课题。分子生物学和基因工程技术的发展,为或得和应用抗菌肽提供了全新的途径。关于抗菌肽基因工程研究的报道甚多,而且不论是原核表达系统还是真核表达系统都可以成功表达目的肽。
然而,原核生物基因工程技术可实现抗菌肽的高效表达。而且可以根据对抗菌肽的认识,通过蛋白质工程技术对现有抗菌肽进行设计改造。从而,获得广谱、高活性、稳定并且适合临床应用的抗菌肽。
抗菌肽的原核生物高效表达的本身也有其需要解决的问题。例如,高效表达的高活性抗菌肽对宿主菌具有强烈的杀伤作用,而且表达出来的抗菌肽产物易被细菌蛋白酶降解。这些问题都对抗菌肽的高水平表达造成了障碍,在基因工程设计的初期就需要考虑到并且解决的。例如, Chen等(2009)通过设计将cecropinA与ceropinD的不同片段结合在一起,实现了天蚕素CAD的高效表达。随着人们对抗菌肽认识的进一步深入,抗菌肽的基因工程将越来越显示出优势。
抗菌肽作为饲料添加剂的应用
近年来,动物抗菌肽作传统抗菌剂代用品的应用受到了人们高度的重视。抗菌肽的抗菌等生物活性使其可成为用于促进畜禽生长、保健和治疗疾病的无毒副作用、无残留、无致细菌耐药性的一类环保型制剂。抗菌肽杀菌机制与传统抗生素不同,病原菌通常不易对抗菌肽产生耐药性。因此,有着广阔的开发应用前景。如能实现抗菌肽的工业化生产,降低成本,则抗菌肽可作为畜禽饲料添加剂取代或部分取代目前饲喂动物所用的抗生素,减少抗生素对动物体的危害。
据欧盟第70/524号令的规定,传统抗生素在2006年以后全部禁止作为饲料添加剂用。为了保证动物源性食品安全,农业部制定了《食品动物禁用的兽药及其它化合物清单》,并于2002年3月5日以农业部农牧发[2002]1号文件发布。传统抗生素作饲料添加剂的使用即将成为历史。应用无毒无公害的新型抗菌剂已成为今后的发展趋势。